

Содержание
В законХарди-Вайнберг, также называемый принципом Харди-Вайнберга или равновесием, состоит из математической теоремы, которая описывает гипотетическую диплоидную популяцию с половым воспроизводством, которое не развивается — частоты аллелей не меняются от поколения к поколению.
Этот принцип предполагает пять условий, необходимых для того, чтобы популяция оставалась постоянной: отсутствие потока генов, отсутствие мутаций, случайное спаривание, отсутствие естественного отбора и бесконечно большой размер популяции. Таким образом, в отсутствие этих сил популяция остается в равновесии.
![]()
Когда любое из предыдущих предположений не выполняется, происходит изменение. По этой причине естественный отбор, мутации, миграции и дрейф генов — четыре механизма эволюции.
Согласно этой модели, когда частоты аллелей популяции равны п а также какие, частоты генотипов будут п2, 2pq а также какие2.
Мы можем применить равновесие Харди-Вайнберга при вычислении частот определенных представляющих интерес аллелей, например, для оценки доли гетерозигот в человеческой популяции. Мы также можем проверить, находится ли популяция в равновесии, и предложить гипотезы о том, что на эту популяцию действуют силы.
Указатель статей
- 1 Историческая перспектива
- 2 Популяционная генетика
- 3 Что такое равновесие Харди-Вайнберга?
- 4 Пример
- 4.1 Первое поколение мышей
- 4.2 Второе поколение мышей
- 5 предположений о равновесии Харди-Вайнберга
- 5.1 Население бесконечно велико
- 5.2 Нет потока генов
- 5.3 Нет мутаций
- 5.4 Случайная вязка
- 5.5 Без выбора
- 6 Решенные проблемы
- 6.1 Частота носителей фенилкетонурии
- 6.2 Ответ
- 6.3 Находится ли следующая популяция в равновесии Харди-Вайнберга?
- 6.4 Популяция бабочек
- 7 ссылки
Содержание
В законХарди-Вайнберг, также называемый принципом Харди-Вайнберга или равновесием, состоит из математической теоремы, которая описывает гипотетическую диплоидную популяцию с половым воспроизводством, которое не развивается — частоты аллелей не меняются от поколения к поколению.
Этот принцип предполагает пять условий, необходимых для того, чтобы популяция оставалась постоянной: отсутствие потока генов, отсутствие мутаций, случайное спаривание, отсутствие естественного отбора и бесконечно большой размер популяции. Таким образом, в отсутствие этих сил популяция остается в равновесии.
![]()
Когда любое из вышеуказанных предположений не выполняется, происходят изменения. По этой причине естественный отбор, мутации, миграции и генетический дрейф — четыре механизма эволюции.
Согласно этой модели, когда частоты аллелей популяции п Y какие, частоты генотипов будут п2, 2pq Y какие2.
Мы можем применить равновесие Харди-Вайнберга при вычислении частот определенных интересующих аллелей, например, для оценки доли гетерозигот в человеческой популяции. Мы также можем проверить, находится ли популяция в равновесии, и предложить гипотезы о том, что силы действуют на эту популяцию.
Второе поколение мышей
Теперь представьте, что эти зиготы развиваются и становятся взрослыми мышами, которые снова будут производить гаметы. Ожидаем ли мы, что частоты аллелей будут такими же или отличаться от частот предыдущего поколения?
Генотип AA будет производить 36% гамет, в то время как гетерозиготы будут производить 48% гамет, а генотип аа 16%.
Чтобы вычислить новую частоту аллеля, мы складываем частоту гомозиготного плюс половина гетерозиготного, как показано ниже:
Частота аллелей К: 0,36 + ½ (0,48) = 0,6.
Частота аллелей к: 0,16 + ½ (0,48) = 0,4.
Если мы сравним их с исходными частотами, то поймем, что они идентичны. Следовательно, согласно концепции эволюции, поскольку частота аллелей не меняется от поколения к поколению, популяция находится в равновесии — она не развивается.
Основы равновесия
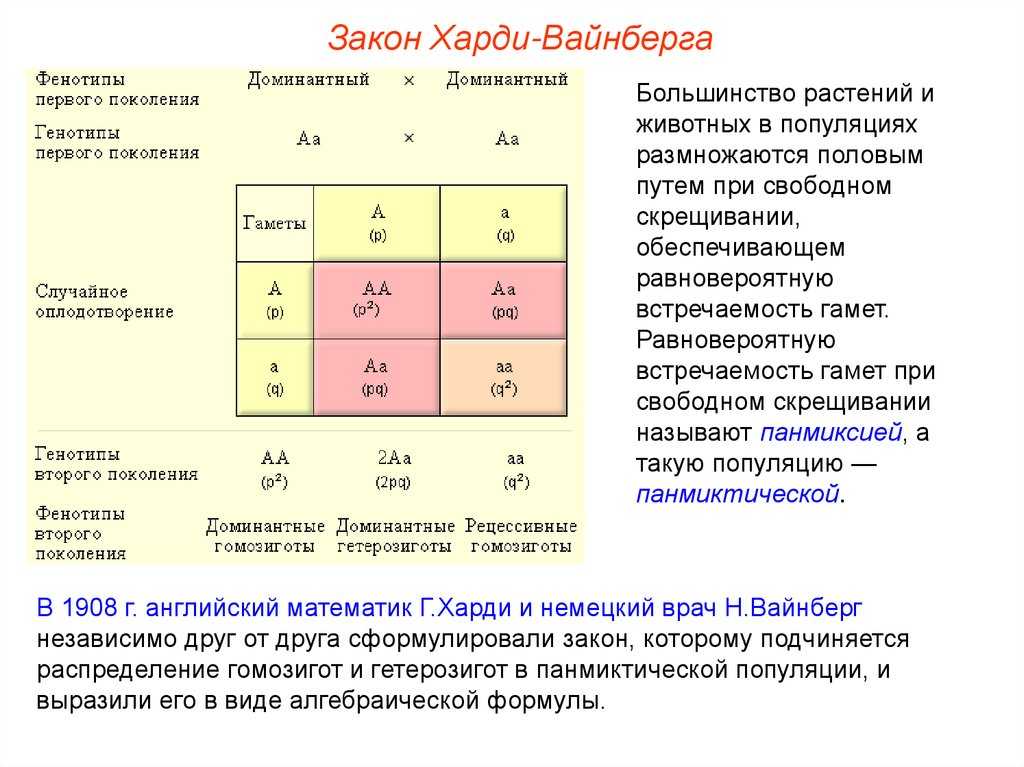
Теорема Харди — Вайнберга имеет дело с менделевской теорией в контексте популяций диплоидных, размножающихся половым путём, особей. Учитывая набор предположений, эта работа гласит:
![]()
- Частоты аллелей в совокупности организмов одного вида не будут меняться на протяжении поколений.
- Если частоты популяционных генотипов с двумя генами в локусе равны p и q, то ожидаемые признаки — p ², 2 pq и q ². Это частотное распределение не изменится для потомков, когда видовое население находится в равновесии Харди-Вайнберга. Например, локусы в конкретной популяциях p и q соответствует аллелям A и a, тогда частота генотипа AA = p ², Aa = 2 pq и aa = q ². Если в локусе только два аллеля, то по необходимости p + q равно единице. Закон Харди — Вайнберга и его математическое выражение (в виде уравнения) выглядит следующим образом: (p + q) ² = p ² + 2 pq + q ² = 1. Применение этой теоремы возможно и к локусам с более чем двумя версиями генов. В этом случае ожидаемые частоты генотипов определяются мультиномиальным разложением для всех k аллелей, выделяющихся в популяции: (p1 + p2 + p3 +… + pk) ².
Выводы из теоремы Харди — Вайнберга применимы только тогда, когда совокупность соответствует следующим допущениям:
![]()
- Естественный отбор не действует на данный локус. То есть нет постоянных различий в вероятностях выживания или размножения между генотипами.
- Ни мутация (происхождение новых аллелей), ни миграция (перемещение особей или их генов в популяцию или из неё) не привносят в видовое сообщество новых генотипов.
- Размер популяции бесконечен. Это означает, что генетический дрейф не вызывает случайных изменений в частотах аллелей из-за ошибки выборки от одного поколения к следующему. Очевидно, что все естественные видовые совокупности конечны и, следовательно, подвержены дрейфу. Однако ожидается, что в малых популяциях такой эффект будет более выраженным.
- Скрещивание людей, по отношению к рассматриваемому локусу, происходит случайным образом. Хотя неслучайное спаривание не меняет версий генов на протяжении поколений, если выполняются другие условия. Например, оно может генерировать отклонения от ожидаемых частот генотипа или подготовить почву для естественного отбора, чтобы вызвать эволюционные изменения.
Если версии генов отклоняются от формулировки закона Харди — Вайнберга, то для того, чтобы привести их в равновесные пропорции, требуется только одно поколение случайных спариваний. Но только при условии, что вышеупомянутые предположения верны, а частоты аллелей равны у мужчин и у женщин (или что особи являются гермафродитами), и этот локус аутосомен.
Учитывая эти условия, легко получить ожидаемые частоты генотипа Харди — Вайнберга, если подумать о случайном спаривании с точки зрения вероятности создания каждого набора версий генов посредством случайного объединения гамет в зиготы. Поскольку существует два способа формирования гетерозиготных генотипов (А или а яйцеклетка и а или А сперматозоид), просуммировать вероятности этих двух типов союзов и прийти к ожидаемой частоте, согласно формуле Харди — Вайнберга.
Историческая перспектива
Принцип Харди-Вайнберга родился в 1908 году и обязан своим именем своим ученым Г.Х. Харди и В. Вайнберг, которые самостоятельно пришли к одним и тем же выводам.
До этого другой биолог по имени Удни Йоль занимался этой проблемой в 1902 году. Йоль начал с набора генов, в которых частоты обоих аллелей были 0,5 и 0,5. Биолог показал, что частоты сохранялись в течение следующих поколений.
Хотя Йоль пришел к выводу, что частоты аллелей можно поддерживать стабильными, их интерпретация была слишком буквальной. Он полагал, что единственное состояние равновесия было найдено, когда частоты соответствовали значению 0,5..
Йоль горячо обсуждал результаты своего романа с Р.С. Пуннетт — широко известен в области генетики благодаря изобретению знаменитой «коробки Пуннетта». Хотя Паннетт знал, что Йоль был неправ, он не нашел математического способа доказать это..
Поэтому Паннетт связался со своим математическим другом Харди, которому удалось немедленно решить его, повторив вычисления, используя общие переменные, а не фиксированное значение 0,5, как это сделал Йоль..
пример
В популяции особи, составляющие ее, скрещиваются, чтобы дать потомство. В общем, мы можем указать на наиболее важные аспекты этого репродуктивного цикла: производство гамет, их слияние, чтобы дать начало зиготе, и развитие эмбриона, чтобы дать начало новому поколению.




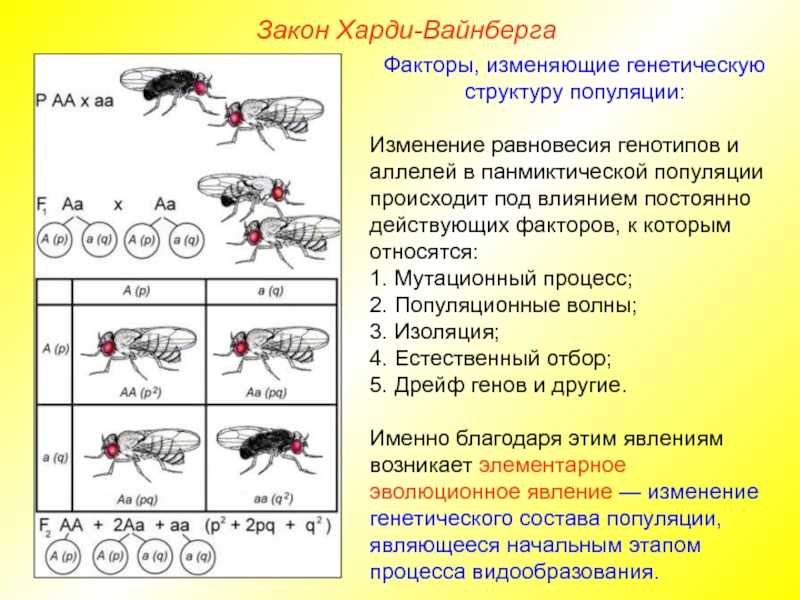



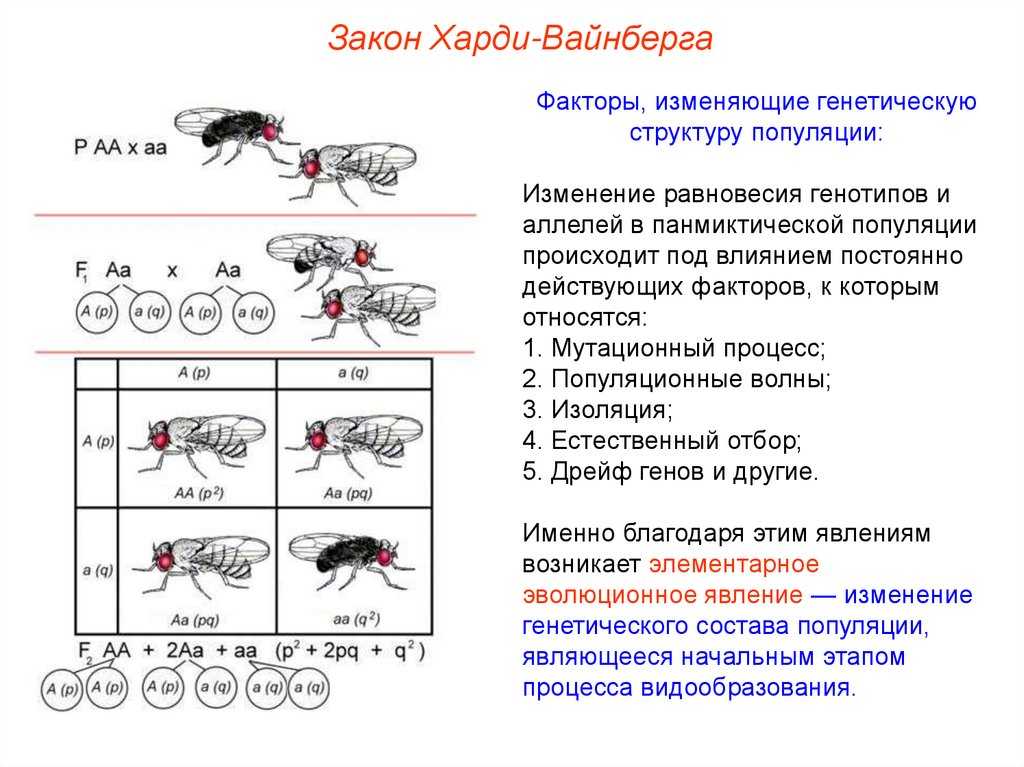
Представим, что мы можем проследить менделевский генный процесс в упомянутых событиях. Мы делаем это, потому что хотим знать, будет ли частота аллеля или генотипа увеличиваться или уменьшаться и почему.
Чтобы понять, как изменяются частоты генов и аллелей в популяции, мы проследим за производством гамет у группы мышей. В нашем гипотетическом примере спаривание происходит случайным образом, когда все сперматозоиды и яйца перемешиваются случайным образом.
В случае мышей это предположение неверно и является упрощением для облегчения вычислений. Однако у некоторых групп животных, таких как некоторые иглокожие и другие водные организмы, гаметы выбрасываются и сталкиваются случайным образом.
Ответить
Чтобы применить уравнение Харди-Вайнберга, мы должны предположить, что выбор партнера не связан с геном, связанным с патологией, и нет инбридинга.
Кроме того, мы предполагаем, что в Соединенных Штатах нет миграционных явлений, нет новых мутаций фенилкетонурии, а вероятность воспроизводства и выживания одинакова для разных генотипов.
Если указанные выше условия верны, мы можем использовать уравнение Харди-Вайнберга для выполнения вычислений, относящихся к проблеме.
Мы знаем, что на каждые 10 000 рождений приходится один случай заболевания, поэтому какие2 = 0,0001, а частота рецессивного аллеля будет квадратным корнем из этого значения: 0,01.
Как p = 1 — какие, мы должны п составляет 0,99. Теперь у нас есть частота обоих аллелей: 0,01 и 0,99. Частота носителей относится к частоте гетерозигот, которая рассчитывается как 2pq. Таким образом, 2pq = 2 х 0,99 х 0,01 = 0,0198.
Это составляет примерно 2% населения. Помните, что это только приблизительный результат.
Первое поколение мышей
Теперь давайте сосредоточим наше внимание на конкретном локусе с двумя аллелями: К а также к. Согласно закону, провозглашенному Грегором Менделем, каждая гамета получает аллель из локуса A
Предположим, что 60% яйцеклеток и сперматозоидов получают аллель. К, а остальные 40% получили аллель к.
Следовательно, частота аллеля К составляет 0,6 и аллеля к составляет 0,4. Эта группа гамет будет найдена случайным образом, чтобы дать начало зиготе. Какова вероятность того, что они сформируют каждый из трех возможных генотипов? Для этого мы должны умножить вероятности следующим образом:
Генотип AA: 0,6 х 0,6 = 0,36.
Генотип Аа: 0,6 х 0,4 = 0,24. В случае гетерозиготы она может возникать в двух формах. Первое, что сперма несет аллель К и яйцеклетка аллель к, или наоборот, сперма к и яйцеклетка К. Поэтому прибавляем 0,24 + 0,24 = 0,48.
Генотип аа: 0,4 х 0,4 = 0,16.
Историческая перспектива
Принцип Харди-Вайнберга родился в 1908 году и обязан своим названием своим ученым Г. Харди и В. Вайнберг, которые независимо пришли к тем же выводам.
До этого другой биолог по имени Удный Юл в 1902 году занялся этой проблемой. Юл начал с набора генов, в которых частоты обоих аллелей были 0,5 и 0,5. Биолог показал, что частота сохраняется в следующих поколениях.
Хотя Юл пришел к выводу, что частоты аллелей можно поддерживать стабильными, его интерпретация была слишком буквальной. Он считал, что единственное состояние равновесия было найдено, когда частоты соответствовали значению 0,5.
Юл горячо обсуждала свои новые открытия с Р.К. Паннетт — широко известен в области генетики изобретением знаменитой «площади Паннета». Хотя Паннетт знал, что Юл был неправ, он не нашел математического способа доказать это.
По этой причине Паннетт связался со своим другом-математиком Харди, который сумел решить ее немедленно, повторив вычисления с использованием общих переменных, а не фиксированного значения 0,5, как это сделал Юл.
Применение закона Харди-Вайнберга
В большинстве случаев частоту аллелей и генотипов вычисляют, взяв за основу частоту гомозиготных особей по рецессивному аллелю. Это единственный генотип, который распознается по фенотипическому выражению. Отличить доминантные гомозиготы от гетерозигот часто не представляется возможным, поэтому их вычисляют, пользуясь уравнением Харди-Вайнберга.
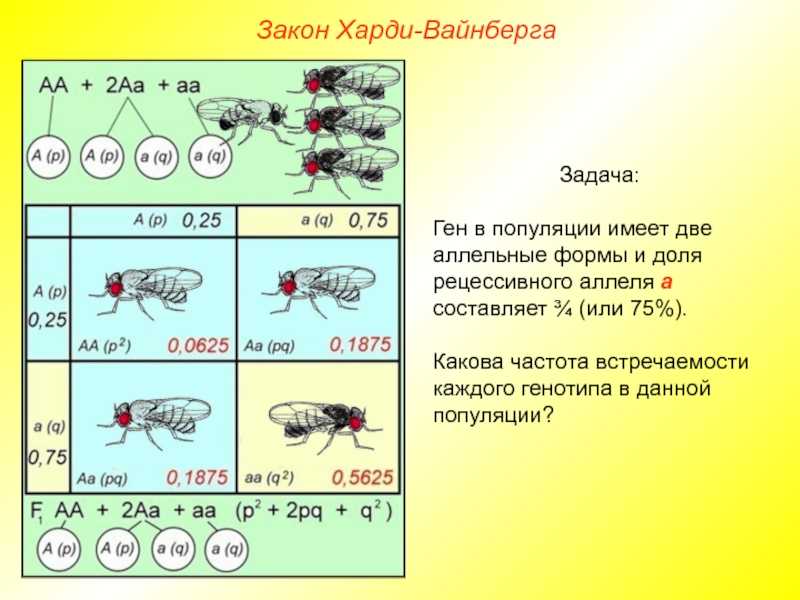
Рассмотрим пример использования закона Харди-Вайнберга. Допустим, в гипотетической популяции людей присутствуют только два аллеля цвета глаз — карий и голубой. Карий цвет определяется доминантным (A) аллелем гена, голубой — рецессивным (a). Пусть кареглазых людей будет 75% (или в долях 0,75), а голубоглазых 25% (или 0,25). Требуется определить, 1) долю гетерозигот и доминантных гомозигот, а также 2) частоту аллелей в популяции.
Если доля рецессивных гомозигот составляет 0,25, то доля рецессивного аллеля находится как квадратный корень из этого числа (исходя из формулы p 2 + 2pq + q 2 = 1, где q 2 — частота рецессивных гомозигот, а q — частота рецессивного аллеля), т. е. будет 0,5 (или 50%). Поскольку в гипотетической популяции людей только два аллеля, то сумма их долей составит единицу: p + q = 1. Отсюда находим долю доминантного аллеля: p = 1 — 0,5 = 0,5. Таким образом, частота обоих аллелей составляет по 50%. Мы ответили на второй вопрос.
Частота гетерозигот составляет 2pq. В данном случае 2 * 0,5 * 0,5 = 0,5. Отсюда следует, что из 75% кареглазых людей 50% являются гетерозиготами. Тогда на долю доминантных гомозигот остается 25%. Мы ответили на первый вопрос задачи.
Другой пример применения закона Харди-Вайнберга. Такое заболевание человека как муковисцидоз встречается только у рецессивных гомозигот. Частота заболевания составляет примерно 1 больной на 2500 человек. Это значит, что 4 человека из 10000 являются гомозиготами, что составляет в долях единицы 0,0004. Таким образом, q 2 = 0,0004. Извлекая квадратный корень, находим частоту рецессивного аллеля: q = 0,02 (или 2 %). Частота доминантного аллеля будет равна p = 1 — 0,02 = 0,98. Частота гетерозигот: 2pq = 2 · 0,98 · 0,02 = 0,039 (или 3,9 %). Значение частоты гетерозигот позволяет оценить количество патогенных генов, находящихся в скрытом состоянии.
Такие вычисления показывают, что, несмотря на малое число особей с гомозиготным рецессивным генотипом, частота рецессивного аллеля в популяциях достаточно велика за счет его нахождения в генотипах гетерозигот (носителей).
Нет выбора
Не существует разницы в репродуктивном успехе людей с разными генотипами, которые могут изменить частоты аллелей в популяции.
Другими словами, в гипотетической популяции все генотипы имеют одинаковую вероятность воспроизводства и выживания.
Когда популяция не соответствует этим пяти условиям, результатом является эволюция. По логике, естественные популяции не соответствуют этим предположениям. Таким образом, модель Харди-Вайнберга используется в качестве нулевой гипотезы, которая позволяет нам делать приблизительные оценки частот генов и аллелей.
Помимо отсутствия этих пяти условий, есть и другие возможные причины, по которым население не сбалансировано.
Одно из них происходит, когда места связаны с полом или явлениями искажения при сегрегации или мейотический драйв (когда каждая копия гена или хромосомы не передается с равной вероятностью следующему поколению).
Каков баланс Харди-Вайнберга?
Равновесие Харди-Вайнберга — это нулевая модель, которая позволяет нам определять поведение частот генов и аллелей на протяжении поколений. Другими словами, это модель, которая описывает поведение генов в популяциях в ряде конкретных условий..










обозначение
В теореме Харди-Вайнберма аллельная частота (доминантный аллель) представлен буквой р, в то время как аллельная частота в (рецессивный аллель) представлен буквой Q.
Ожидаемые генотипические частоты р2, 2рд и Q2, для доминантного гомозиготы (А.А.), гетерозиготный (Aa) и рецессивный гомозигот (аа) соответственно.
Если в этом локусе только два аллеля, сумма частот двух аллелей обязательно должна быть равна 1 (p + q = 1). Биноминальное расширение (p + q)2 представляют генотипические частоты р2 + 2pq + q2 = 1.
Проблемы решены
Частота фенилкетонурии носителей
По оценкам, в Соединенных Штатах у одного из 10000 новорожденных есть состояние, которое называется фенилкетонурия..
Это расстройство выражается только у рецессивных гомозигот при нарушении обмена веществ. Зная эти данные, какова частота носителей заболевания в популяции??
ответ
Чтобы применить уравнение Харди-Вайнберга, мы должны предположить, что выбор партнера не связан с геном, связанным с патологией, и нет инбридинга..
Кроме того, мы предполагаем, что в Соединенных Штатах нет миграционных явлений, нет новых мутаций фенилкетонурии, а вероятность размножения и выживания одинакова для генотипов..
Если вышеупомянутые условия выполняются, мы можем использовать уравнение Харди-Вайнберга для выполнения расчетов, имеющих отношение к проблеме.
Мы знаем, что заболевание случается каждые 10 000 рождений, поэтому Q2 = 0,0001 и частота рецессивного аллеля будет квадратным корнем из этого значения: 0,01.
в качестве р = 1 — Q, мы должны р Это 0,99. Теперь у нас есть частота обоих аллелей: 0,01 и 0,99. Частота носителей относится к частоте гетерозигот, которая рассчитывается как 2рд. Итак, 2рд = 2 х 0,99 х 0,01 = 0,0198.
Это эквивалентно примерно 2% населения. Напомним, что это только приблизительный результат.
Следующая популяция находится в равновесии Харди-Вайнберга?
Если мы знаем число каждого генотипа в популяции, мы можем заключить, находится ли он в равновесии Харди-Вайнберга. Шаги для решения этого типа проблем следующие:
- Рассчитайте наблюдаемые генотипические частоты (D, H и R)
- Рассчитаем частоты аллелей (р и Q)
p = D + ½ H
q = R + ½ H
- Рассчитать ожидаемые генотипические частоты (р2, 2PQ и Q2)
- Рассчитать ожидаемые числа (р2, 2PQ и Q2), умножая эти значения на общее количество особей
- Сравните ожидаемые цифры с теми, которые наблюдались с помощью теста X2 от Пирсона.
Популяция бабочек
Например, мы хотим проверить, находится ли следующая популяция бабочек в равновесии Харди-Вайнберга: существует 79 особей гомозиготного доминантного генотипа (А.А.) 138 гетерозигот (Aa) и 61 рецессивный гомозигот (аа).
Первым шагом является вычисление наблюдаемых частот. Мы делаем это путем деления числа особей по генотипу на общее число особей:
D = 79/278 = 0,28
H = 138/278 = 0,50
R = 61/278 = 0,22
Чтобы проверить, хорошо ли я сделал это первый шаг, я добавляю все частоты и должен дать 1.
Второй шаг — вычислить частоты аллелей..
р = 0,28 + ½ (0,50) = 0,53
Q = 0,22 + ½ (0,50) = 0,47
С этими данными я могу рассчитать ожидаемые частоты генотипа (р2, 2PQ и Q2)
р2 = 0,28
2pq = 0,50
Q2 = 0,22
Я рассчитываю ожидаемые числа, умножая ожидаемые частоты на количество особей. В этом случае число наблюдаемых и ожидаемых особей одинаково, поэтому я могу сделать вывод, что население находится в равновесии.
Если полученные числа не совпадают, я должен применить вышеупомянутый статистический тест (X2 от Пирсона).
Закон Харди—Вайнберга
Чтобы изменить состав генофонда, требуется нечто большее, чем генетическая рекомбинация.
В научном мире нечасто случается, чтобы разные ученые независимо друг от друга наткнулись на одну и ту же закономерность, но все же таких примеров достаточно, чтобы заставить нас поверить в существование «духа времени». К их числу относится и закон Харди—Вайнберга (известный также как закон генетического равновесия) — одна из основ популяционной генетики. Закон описывает распределение генов в популяции. Представьте себе ген, имеющий два варианта — или, пользуясь научной терминологией, два аллеля. Например, это могут быть гены «низкорослости» и «высокорослости», как в случае менделевского гороха (см. Законы Менделя), или наличие/отсутствие предрасположенности к рождению двойни. Харди и Вайнберг показали, что при свободном скрещивании, отсутствии миграции особей и отсутствии мутаций относительная частота индивидуумов с каждым из этих аллелей будет оставаться в популяции постоянной из поколения в поколение. Другими словами, в популяции не будет дрейфа генов.
Рассмотрим этот закон на простом примере. Назовем два аллеля Х и х. Тогда у особей могут встречаться четыре следующие комбинации этих аллелей: ХХ, хх, хХ и Хх. Если обозначить через p и q частоту встречаемости индивидуумов с аллелями Х и х соответственно, то согласно закону Харди—Вайнберга
![3.2. закон харди - вайнберга и его приложения [1989 фогель ф., мотульски а.г. - генетика человека. в 3-х т. том 1]](https://babymart-ekb.ru/wp-content/uploads/2/b/6/2b6787bdb094a01a6c6795b44f8021d2.jpeg)







где p 2 — частота встречаемости индивидуумов с аллелями ХХ, 2pq — с аллелями Хх или хХ, а q 2 — частота встречаемости индивидуумов с аллелями хх. Эти частоты, при соблюдении сформулированных выше условий, будут оставаться постоянными из поколения в поколение, независимо от изменения количества индивидуумов и от того, насколько велики (или малы) p и q. Этот закон представляет собой модель, используя которую генетики могут количественно определять изменения в распределении генов в популяции, вызванные, например, мутациями или миграцией. Другими словами, этот закон является теоретическим критерием для измерения изменений в распределении генов.
Английский математик, родился в Кранли, графство Суррей. Сын учителя рисования. Изучал математику в Кембриджском и Оксфордском университете. Пожалуй, самую большую известность Харди принесли совместные работы с Джоном Идензором Литлвудом (John Edensor Littlewood, 1885–1977) и позднее с индийским математиком-самоучкой Cриниваса Рамануджаном (Srinivasa Aaiyangar Ramanujan, 1887–1920), который работал клерком в Мадрасе. В 1913 году Рамануджан послал Харди список доказанных им теорем. Признав гениальность юного клерка, Харди пригласил его в Кембридж, и в течение нескольких лет, предшествовавших безвременной смерти Рамануджана, они опубликовали серию блестящих совместных работ.
Немецкий врач, имевший большую частную практику в Штуттгарте. По воспоминаниям современников, помог появиться на свет 3500 младенцам, в том числе по крайней мере 120 парам близнецов. На основании собственных наблюдений над рождением близнецов и переоткрытых генетических законов Менделя пришел к выводу, что предрасположенность к рождению двуяйцевых (неидентичных) близнецов передается по наследству.
Следствия закона
Теорема демонстрирует, что менделевские локусы, сегрегирующие по множественным аллелям в диплоидных популяциях, сохранят предсказуемые уровни генетической изменчивости при отсутствии факторов, которые влияют на структурные изменения генотипов. Основным способом визуализации этих вероятностей является график p ², 2 pq и q ² в зависимости от частот аллелей. Если его нарисовать, то можно увидеть два важных следствия:
- Популяционная гетерозиготность наиболее высокая при выполнении равенства p = q = 0,5.
- Редкие аллели обнаруживаются главным образом у гетерозигот, как и должно быть, учитывая, что q ² намного меньше, чем 2 pq, когда q близко к нулю, и p ² мало, в соотношении с 2 pq, если p стремится к 0.
![]()
Второй пункт приобретает особое значение, если рассмотреть возможность естественного отбора влиять на частоты новых мутаций. Когда популяция соответствует всем другим вероятностям закона, отбор в конечном счёте определит полезный ген, так что все индивидуумы будут гомозиготными по этому аллелю. Первоначальное увеличение частоты редкого, выгодного, доминантного гена происходит быстрее, чем у редкого, рецессивного аллеля.
Новая доминантная мутация характеризуется тем, что она сразу видна при естественном отборе, потому что его влияние на физическую форму проявляется у гетерозигот. Таким образом, Харди продемонстрировал, что доминирование само по себе не изменяет частоты аллелей в локусе, но, как правило, оказывает существенное влияние на эволюционные траектории.
Второе поколение мышей
Теперь давайте представим, что эти зиготы развиваются и становятся взрослыми мышами, которые снова будут производить гаметы. Ожидаем ли мы, что частоты аллелей будут такими же или отличаться от частот предыдущего поколения?
Генотип AA будет производить 36% гамет, в то время как гетерозиготы будут производить 48% гамет, а генотип аа 16%.
Чтобы рассчитать новую частоту аллеля, мы складываем частоту гомозиготного плюс половина гетерозиготного, как показано ниже:
Частота аллелей К: 0,36 + ½ (0,48) = 0,6.
Частота аллелей к: 0,16 + ½ (0,48) = 0,4.
Если мы сравним их с исходными частотами, то поймем, что они идентичны. Следовательно, согласно концепции эволюции, поскольку нет изменений в частотах аллелей на протяжении поколений, популяция находится в равновесии — она не развивается.
Популяционная генетика
Популяционная генетика направлена на изучение сил, которые приводят к изменению частот аллелей в популяциях, объединяя теорию эволюции Чарльза Дарвина посредством естественного отбора и менделевской генетики. В настоящее время его принципы обеспечивают теоретическую основу для понимания многих аспектов эволюционной биологии..
Одной из важнейших идей популяционной генетики является взаимосвязь между изменениями относительного обилия признаков и изменениями относительного обилия аллелей, которые их регулируют, что объясняется принципом Харди-Вайнберга. Фактически, эта теорема обеспечивает концептуальную основу для популяционной генетики..
В свете популяционной генетики концепция эволюции заключается в следующем: изменение частот аллелей на протяжении поколений. Когда нет изменений, нет эволюции.